研究内容
1. シアル酸分子種の生物学的役割に関する研究
私たちの細胞は例外なく糖鎖で覆われており、これらの糖鎖の末端の多くはシアル酸と呼ばれる酸性の糖で占められています。このため、シアル酸は細胞の分子認識に様々な形で関わっています。例えば、インフルエンザウイルスは細胞表面のシアル酸を含む糖鎖と結合することでその感染を開始し、シアル酸を切断することで感染細胞から出芽します。これは、シアル酸が細胞の一番外側に存在すること、また、マイナスのチャージをもつなどの性質によります。シアル酸はノイラミン酸骨格という構造をもつ酸性糖の総称であり、非常に多くの分子修飾を受けます。細胞は様々な分子を認識し、それに応じた応答を行うわけですが、多様な構造をもつシアル酸による糖鎖修飾は、これら複雑な分子間認識を可能にするために制御されていると考えられています。私たちは、このシアル酸分子を介した分子認識の機能的な意義を主に免疫系において解析しています。
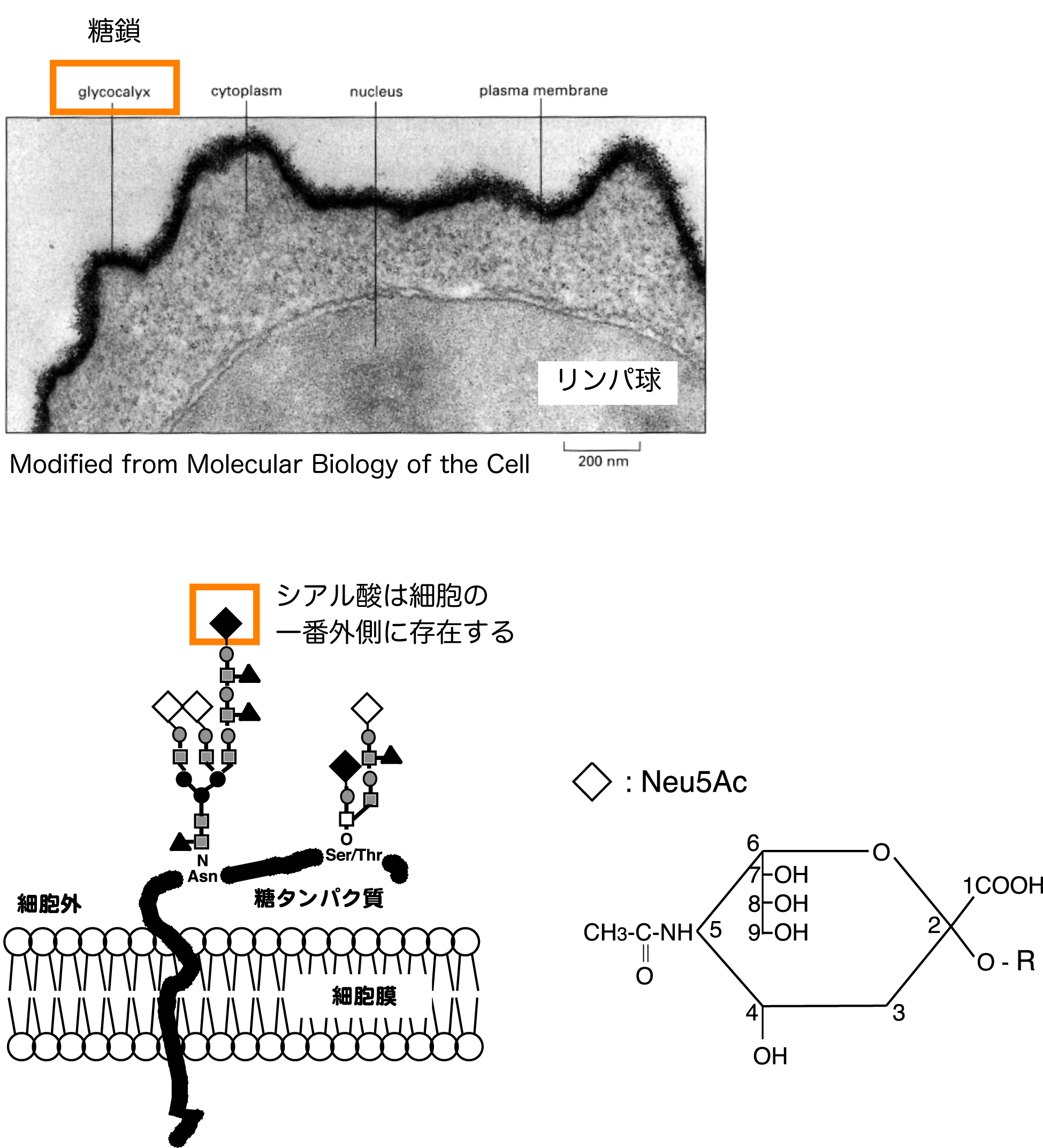
マウスリンパ球細胞は、Neu5Gcを主要なシアル酸として発現しています。私たちは、抗原からの刺激によりB細胞を活性化することでNeu5Gcの発現が抑制されること、このNeu5Gcの抑制はCD22と呼ばれるB細胞活性化抑制能をもつ分子のリガンド(標的分子)の発現を低下させることを明らかにしました。また、B細胞は抗原刺激に応答して抗体を産生しますが、このための活性化の場である胚中心を特異的に染色する単クローン抗体GL7が、活性化リンパ球(T細胞でもこの反応は起こる)特異的なNeu5Acの発現を認識していることを明らかにしました。私たちはNeu5Gcの発現を動物個体レベルで変化させた遺伝子欠損マウス、トランスジェニックマウスなどの系を用いて、シアル酸分子種の違いがもたらす免疫学的な応答を明らかにすべく研究を続けています。
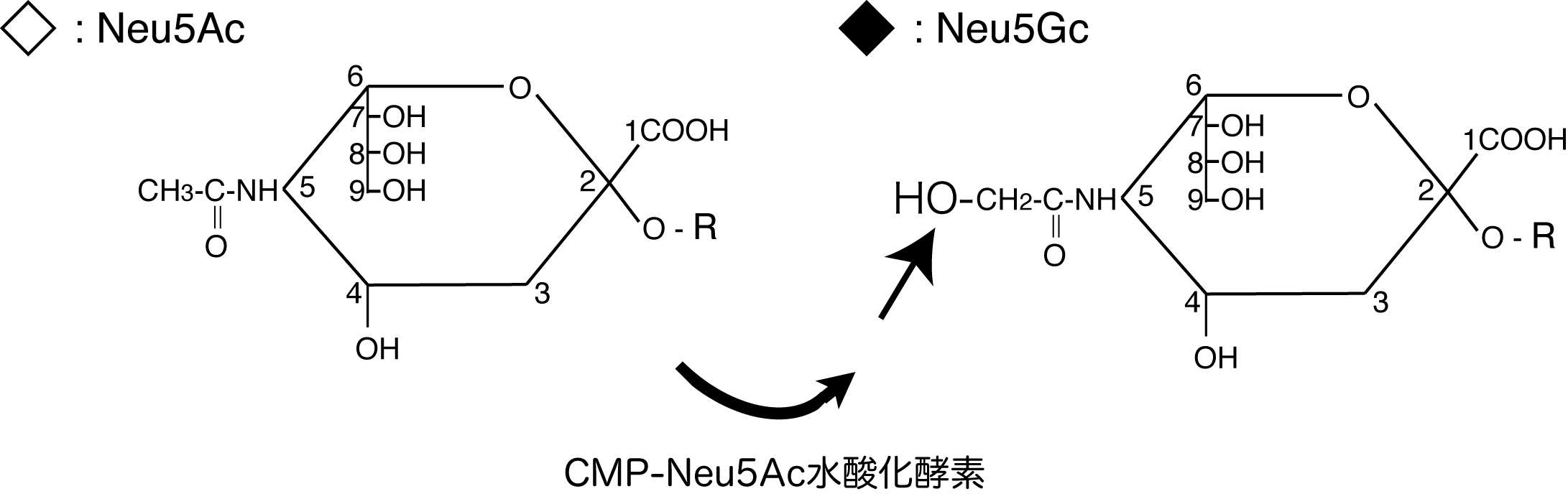
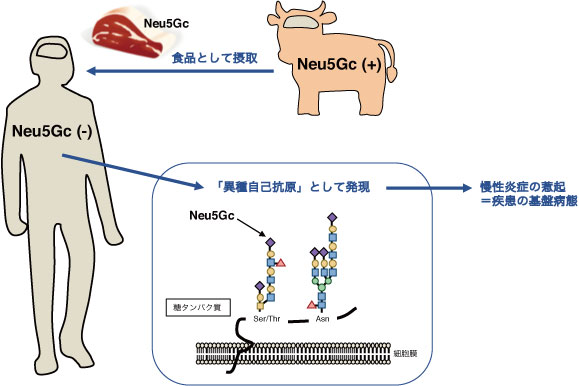
一方、ヒトはNeu5Gcの生合成酵素であるCMP-Neu5Ac水酸化酵素を欠失しています。このCMP-Neu5Ac水酸化酵素の遺伝子は、ヒトで欠損しているものの、現存生物の中でヒトと最近縁であり遺伝子レベルでも98%同じとされるチンパンジーやボノボでは欠損しておらず、Neu5Gcの発現はヒトとその他の動物を分ける違いであることが分かりました。シアル酸は体中の細胞で発現しているため、この酵素の欠損は全身において影響を及ぼす可能性があリます。ヒト免疫系から考えるとNeu5Gcは異種性の非自己抗原となりますが、実際には食事から取り込まれ自己抗原として後天的に発現するという他に類をみない「異種自己抗原」であり、しかもNeu5Gcが抗原性をもつことで慢性の自己応答性抗原となることが報告されています。私たちは、ヒトで特異的に起こる異種自己抗原Neu5Gcに対する免疫応答機構を明らかにするとともに、これがヒトの炎症性疾患に対し及ぼす影響を、Neu5Gc欠損マウスを用いて研究しています。この異種自己抗原応答を人為的にコントロールすることで、現在も改良が求められている、よりヒトの状態を模倣した疾患モデルマウスを構築できると考えています。今後ヒトの治療を念頭に置いたマウスモデルでの実験において、Neu5Gc欠損を標準背景とすることが必要になると考えています。
2. スフィンゴ脂質が関与するシグナル伝達機構に関する研究
細胞膜は様々な成分により構成されており、その一つにスフィンゴ脂質と呼ばれる脂質があります。この脂質は他の膜成分であるグリセロリン脂質と比べるとその機能はほとんど明らかにされていませんでしたが、近年、スフィンゴ脂質の生理的意義に関する議論が展開され、その重要性が徐々に明らかになりつつあります。当研究室では、いくつかのスフィンゴ脂質に関する試薬を中心に研究を進めています。
1) 近年、様々な免疫抑制剤の登場が臓器移植や自己免疫疾患などの治療に大きな貢献をもたらしてきました。しかし、これらの免疫抑制剤は同時に、重篤な腎障害などの副作用を引き起こす事が知られているため、今ではこれまでとは異なった作用機序を有する免疫抑制剤の開発が望まれています。この様な背景の中、冬虫夏草の一種であるタイワンツクツクホウシを宿主とするIsalia sinclairii菌から強い免疫抑制活性を持つ物質が単離され、ISP-1 (immunosupressant product-1)と名付けられました。構造解析から、ISP-1は従来より抗菌活性物質として知られているmyriocin, thermozymocidinと同一のものでしたが、免疫抑制作用としては初めての知見でした。また、その構造はスフィンゴ脂質に類似しており、構造上類似点の乏しい他の免疫抑制剤とは異なる作用機序が予想されました。そこで、ISP-1による免疫抑制作用の解析を続けたところ、スフィンゴ脂質の生合成阻害によるスフィンゴ脂質の減少が非常に重要な役割を持つことが明らかにされました。このISP-1をリード化合物として、これまでにないタイプの免疫抑制剤であるFTY720(フィンゴリモド)が開発されました。FTY720は2010年に自己免疫疾患の一つである多発性硬化症の世界初の経口薬として米国及びロシアにおいて承認されています。これまでに当研究室では、酵母においてISP-1耐性遺伝子の一つとしてYPK1を単離し、Ypk1がスフィンゴ脂質を介するシグナル伝達系の下流に存在している事を明らかにしました。Ypk1はISP-1耐性のみならず、細胞増殖、エンドサイトーシス、アクチン形成を正に制御していることが知られていますが、これらの制御機構の詳細は明らかになっていません。私たちはこれらのシグナル伝達系を明らかにすべく解析を行っています。また、当研究室ではYpk1が栄養源の一つである窒素源の飢餓時に、オートファジー及びエンドサイトーシスの二つの経路によって液胞へと輸送されて選択的に分解されることを明らかにしました。私たちが発見したこの経路はこれまで知られていない新規の経路であり、これに関してもその詳細を明らかにしようとしています。
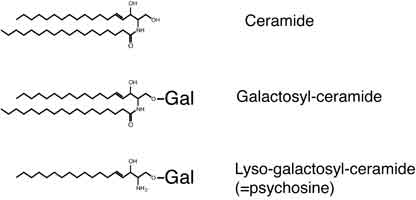
2) 脂肪酸を1本のみもつリゾ型スフィンゴ脂質は、2本の脂肪酸をもつ通常のスフィンゴ脂質とは異なる物性を膜に与えるだけでなく、これら自身が様々な生理活性を有しています。私たちはこれまでに、リゾ型の生体内糖脂質であるサイコシンは細胞周期のうち細胞質分裂を特異的に阻害することで、細胞の多核化、巨大化を誘導することを解明してきました。また、サイコシン処理した細胞はトランスゴルジネットワーク様の細胞内小胞の再分布を促進することを示しました。しかしながら、細胞分裂の最終段階である細胞質分裂のみを阻害し、細胞周期を停止させないという非常に興味深いサイコシンの標的作用機序に関しては、いまだ謎に包まれています。そこで私たちは、サイコシン標的因子、さらには、サイコシン以外のリゾスフィンゴ脂質の作用機序を解明するべく研究を進めています。